
Dispar
The Online Journal of Lepidoptera
ISSN 2056-9246
The Online Journal of Lepidoptera
ISSN 2056-9246
Abstract: It has been speculated that climate change may reduce butterfly numbers as a result of asynchrony of butterfly species with their essential environmental resources. Additionally climate change may induce them to attempt to breed when conditions are initially conducive to adult ovipositing but ultimately sub-critical for full larval development. Both of these phenomena would be detrimental to the long term survival of the species and raise questions about the evolutionary value of responding to climate change in the way that has been documented by various authors, such as attempting to breed both earlier and later in the year. An observation of an 'extra brood', resulting in larvae of Large White Pieris brassicae very late in the year, suggests that, just as asynchrony between plants and butterflies may prove challenging to the butterflies' survival, asynchrony with the autecology of their parasitoids can assist survival. If further studies confirm this finding, it would explain the value of a genetic component giving a predilection towards attempting additional broods despite the potential pitfalls as essential environmental resources dwindle.
A recent study by Hans Van Dyck et al. (2015) speculates that, prompted by environmental cues (that result from global warming), the Wall Brown Lasiommata megera has recently begun to produce an autumn brood in regions that were previously too cold for this. Though conditions induce the adult butterfly to fly, mate and lay eggs in the early autumn, it is suggested that subsequent larval development is often impossible; with mortality so great that subsequent population size is catastrophically weakened. Such responses to climate change are considered to be part of the plastic response that genetics confer on many species and the outcomes can be both beneficial and harmful.
When the outcome is plainly harmful, it is hard to conceive of a reason for such a response being retained within the species' genome. This observation suggests how such a response may be a useful trait. Conceivably, though it risks incomplete larval development, the insect may be operating in conditions where its parasites cannot. Consequent losses through incomplete development can be more than offset by exploiting an opportunity to avoid parasitism (which would otherwise cause greater mortality). If this is so, it provides some evidence for Parmesan's (2006) assertion that: "For many species, the primary impact of climate change may be mediated through effects on synchrony with that species' food and habitat resources". It may provide evidence of genetic responses to climate change as proposed by Skelly and Freidenburg (2010).
Observation of six Large White Pieris brassicae larvae climbing the French windows of a house and attempting to pupate in late October 2014, revealed that all were parasitized by a Cotesia parasitoid wasp whose cocoons were visible near the larva. During November, numbers of larvae were still climbing the windows but it was noted that a few pupated successfully (without any parasitoid emerging from the larvae).
In early December larvae apparently achieved pupation (without any parasitoids emerging) more frequently. Consequently on December 10th all the pupae and Cotesia cocoons that could be found adhered to a north east facing wall and an adjacent fence were counted. The positions of these pupae and parasitoid cocoons were marked to differentiate between those that attempted pupation before 10th December and those after.
This thorough count also revealed one larva that had died during the transition to pupa and four pupae with the cocoons of a Tachinid fly beside them.
Mid-December 2014 was mild and larvae continued to feed and develop on garden brassicae; but as we approached the shortest day, it was evident that the lowering temperatures were resulting in lengthy periods of larval inactivity. Consequently, on 18th December, all final instar sized larvae were collected from their brassicae and brought into an unheated, lean to outbuilding; partly to protect against forecast frosts but also to be certain of the fate of these larvae when it came to pupation. (In fact, over the next three weeks, though ground frosts occurred, the air temperature always stayed slightly above freezing).
Smaller larvae (which had been left outside) continued to feed periodically and to develop slowly. On 2nd January 2015, six more of these larvae had reached final instar size and were brought into the unheated outbuilding for close observation of their pupation attempt.
The last three outdoor feeding larvae (though still in their penultimate 4th instar) were brought inside on the 7th January 2015 (after which there were five nights with subzero air temperatures whilst these three continued to develop, on cut foodplant, in a slightly warmer environment). The last larva pupated on 27th January 2015.
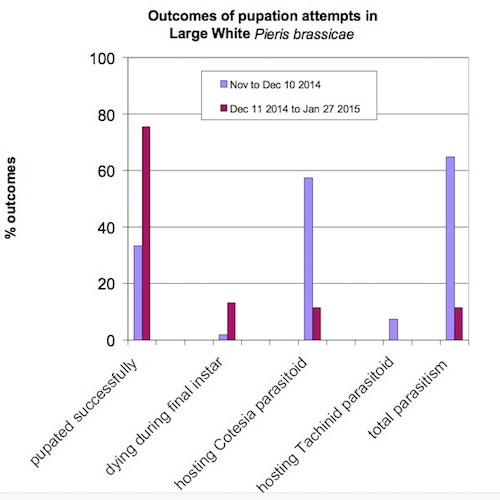
A record of the pupation attempts, for two contiguous periods, is summarised in the tables below: those that attempted pupation between November and December 10th (Table 1) and those that attempted pupation between December 11th and January 27th (Table 2). A comparison of these two periods is then given in Figure 1.
| Number | Percentage | |
| A - Pupated successfully | 18 | 33.33 |
| B - Dying during final instar | 1 | 1.85 |
| C - Hosting Cotesia parasitoid | 31 | 57.41 |
| D - Hosting Tachinid parasitoid | 4 | 7.41 |
| Total parasitism = C+D | 35 | 64.82 |
| Total = A+B+C+D | 54 | 100.00 |
| Number | Percentage | |
| A- Pupated successfully | 46 | 75.41 |
| B - Dying during final instar | 8 | 13.11 |
| C - Hosting Cotesia parasitoid | 7 | 11.48 |
| D - Hosting Tachinid parasitoid | 0 | 0.00 |
| Total parasitism = C+D | 7 | 11.48 |
| Total = A+B+C+D | 61 | 100.00 |
 |
| Figure 1 - A Summary of Pupation Attempts in the Large White Pieris brassicae |
Whilst these results are comparing two intervals of a single observation period, the results are sufficiently marked to suggest that further observations to assess the outcome could be worthwhile.
Two main conclusions emerge from this study. The first is that it is clear that a far larger percentage of larvae reached pupation after 10th December 2014 without evidence of parasitoids. The second is that, of those larvae that were not parasitized, a larger percentage were observed to die in the final instar after December 10th, presumably as a result of unfavourable environmental resources. However, a weakness of the method employed is that the relatively large numbers of densely grouped larvae (living outdoors, before being brought into an outhouse on 18th December 2014) were difficult to observe individually; larvae may have died before that date without being detected.
It appears that in mild early winters, the phenotypic plasticity of the Large White's response to a warmer environment, laying eggs throughout October, is rewarded. There is a greater chance of underdeveloped larvae of this additional brood dying in the cold weather but this is more than offset by the greatly reduced frequency of parasitoid attack.
Whilst in some cases (such as that of the Wall) the cues that prompt some genotypes of a species to attempt an 'extra' brood lead them to inevitable failure (as a result of an inability to cope with subsequent environmental conditions). In other cases there can be advantages of such responses if they allow a species to exploit asynchrony between it and potential parasitoids. If the development of any extra brood continues (albeit more slowly and with greater mortality as a result of unfavourable environmental conditions) it may still prove advantageous, if this extra brood is unencumbered by the huge mortality engendered by a parasitic infestation. A change in the genome to favour alleles that promote broods at the extreme of the critical physiological barrier could be expected in such a case.
